

ABSTRACT
The recently discovered Middle East respiratory syndrome coronavirus (MERS-CoV) continues to infect humans, with high mortality. Specific, highly effective therapeutics and vaccines against the MERS-CoV are urgently needed to save human lives and address the pandemic concerns. We identified three human monoclonal antibodies (MAbs), m336, m337, and m338, targeting the receptor (CD26/DPP4) binding domain (RBD) of the MERS-CoV spike glycoprotein from a very large naïve-antibody library (containing ∼1011 antibodies). They bound with high affinity: equilibrium dissociation constants for the three MAbs were equal to 4.2, 9.3, and 15 nM, respectively, as measured by Biacore for Fabs binding to RBD. The avidity for IgG1 m336, m337, and m338 was even higher: 99, 820, and 560 pM, respectively. The antibodies bound to overlapping epitopes that overlap the receptor binding site on the RBD as suggested by competition experiments and further supported by site-directed mutagenesis of the RBD and a docking model of the m336-RBD complex. The highest-affinity MAb, m336, neutralized both pseudotyped and live MERS-CoV with exceptional potency, 50% neutralization at 0.005 and 0.07 μg/ml, respectively, likely by competing with DPP4 for binding to the S glycoprotein. The exceptionally high neutralization activity of these antibodies and especially m336 suggests that they have great potential for prophylaxis and therapy of MERS-CoV infection in humans and as a tool for development of vaccine immunogens. The rapid identification (within several weeks) of potent MAbs suggests a possibility to use the new large antibody library and related methodology for a quick response to the public threat resulting from emerging coronaviruses.
IMPORTANCE A novel human coronavirus, the Middle East respiratory syndrome coronavirus (MERS-CoV), was found to infect humans with a high mortality rate in 2012, just 1 decade after the appearance of the first highly pathogenic coronavirus, severe acute respiratory syndrome coronavirus (SARS-CoV). There are no effective therapeutics available. It is highly desirable to find an approach for rapidly developing potent therapeutics against MERS-CoV, which not only can be implemented for MERS treatment but also can help to develop a platform strategy to combat future emerging coronaviruses. We report here the identification of human monoclonal antibodies (MAbs) from a large nonimmune antibody library that target MERS-CoV. One of the antibodies, m336, neutralized the virus with exceptional potency. It therefore may have great potential as a candidate therapeutic and as a reagent to facilitate the development of vaccines against MERS-CoV.
INTRODUCTION
In September 2012, a novel human coronavirus, the Middle East respiratory syndrome coronavirus (MERS-CoV), was isolated from a Saudi Arabian patient suffering from severe pneumonia, who later died of respiratory and renal failure (1). Following that, a cluster of 23 laboratory-confirmed cases of MERS-CoV infections were identified between 1 April and 23 May 2013 and investigated, including full-genome sequencing of MERS-CoV from four patients (2). As of 27 March 2014, 206 infections, including 86 deaths, had been reported (http://www.who.int/csr/don/2014_03_27_mers). Although past outbreaks have been contained, more clusters and sporadic cases have originated from multiple sources, indicating a natural animal reservoir and the potential for future outbreaks (3–5). It is also notable that the clinical, epidemiological, and virological features of MERS-CoV are similar to that of the severe acute respiratory syndrome coronavirus (SARS-CoV), which is the first highly pathogenic coronavirus, appearing about 1 decade ago, and caused a global pandemic with more than 800 deaths worldwide within a few months. A number of new coronaviruses have been identified since the outbreak of SARS, and accumulating evidence indicates that coronaviruses are able to rapidly and stably adapt to new host species (6). These findings indicate that SARS-CoV- and MERS-CoV-like novel coronaviruses are highly likely to continue to emerge and pose a serious threat to human health in the near future.
There are currently no effective therapies against MERS-CoV. Recently, a study reported that rhesus macaques treated with alpha 2b interferon (IFN-α2b) and ribavirin, which does not directly target the virus but reduces damaging inflammation of the lung, exhibited reduced MERS-CoV replication and an improved clinical outcome (7). However, the treatment was initiated soon (8 h) after challenge and the disease in the macaques was at best mild to moderate in severity, so whether the drug cocktail would work when faced with severe human disease, which may take longer to develop, remains unclear. Broad-spectrum antivirals have not proven to be effective in the clinic, and in fact there are currently no approved antiviral treatments for any human coronavirus infection. Thus, it is urgent to find an approach to rapidly develop potent therapeutics against MERS-CoV, which not only can be implemented for the treatment of MERS but also can help to develop a platform strategy against emerging coronaviruses in the future.
Monoclonal antibodies (MAbs) are enjoying significant success in the clinic and have been used for the effective treatment of a number of diseases, in particular, cancer and immune disorders (8–11). Although more than 40 MAbs have been approved for clinical use, the humanized MAb palivizumab (Synagis) (12) remains the only MAb approved by the FDA for use against a viral disease. However, an increasing number of MAbs against emerging viruses, as well as against HIV-1 and other viruses, have been developed. Some of these MAbs are promising candidate therapeutics, and some were used on a compassionate basis for prophylaxis of humans exposed to virus (13–15).
We previously developed an MAb, m102.4, against the emerging viruses Hendra virus (HeV) and Nipah virus (NiV), which was administered on a compassionate basis to humans exposed to HeV based on its efficacy in vitro and in animals (16–19). We have also discovered MAbs against SARS-CoV that exhibited potent neutralization of SARS-CoV isolates (20). In this study, we constructed a novel, very large IgM library (containing ∼1011 antibodies) by using B cells from the blood of 40 healthy donors in order to enhance selection of high-affinity antibodies with minimal divergence from their germ line predecessors and engineered a fragment containing the MERS-CoV S protein receptor binding domain (RBD), which is a major MERS-CoV neutralization determinant. This RBD was used as a selecting antigen for panning of the library. A panel of antibodies was identified and characterized. Here we present evidence that three of these antibodies have exceptionally potent neutralization activity against MERS-CoV. These antibodies could be useful for prophylaxis of MERS and treatment of MERS-CoV-infected patients and as reagents to facilitate the development of therapeutics and vaccines as well as to help understand their mechanisms of action.
MATERIALS AND METHODS
Generation, expression, and characterization of MERS-CoV RBD, RBD mutants, and truncated S proteins.
The MERS-CoV S gene segments were synthesized by Genscript (Piscataway, NJ). The plasmid encoding residues 360 to 601 fused with Avi tag was transfected into 293 Freestyle cells (Invitrogen) for transient expression and used for biopanning. Plasmids encoding residues 18 to 725, 18 to 353, 358 to 588, 377 to 588, 377 to 662, and 726 to 1295 fused with the Fc fragment of IgG1 were also transfected into 293T cells (ATCC). Point mutations were generated at specific residues of the MERS-CoV RBD (residues 377 to 588) using the QuikChange II XL site-directed mutagenesis kit (Stratagene). The RBD protein and its mutants were expressed and analyzed as described previously (21).
Generation of a large phage-displayed IgM Fab library, selection, expression, and purification of the RBD-specific Fabs, and conversion to IgG1s.
A large phage display library was constructed by using peripheral blood mononuclear cell (PBMC) cDNA from 40 healthy volunteers as the templates for cloning the expressed antibody gene repertoire. The library was panned using MERS-CoV RBD (residues 360 to 601) conjugated to magnetic beads (Invitrogen). Amplified libraries of 1012 phage-displayed Fabs were incubated with 5, 3, 3, and 1 μg of RBD for 2 h at room temperature during the first, second, third, and fourth rounds of biopanning, respectively. Clones that bound to RBD were identified from the third and fourth rounds of panning by using monoclonal phage enzyme-linked immunosorbent assay (ELISA). The heavy chain variable domain (VH) and light chain variable domain (VL) of these clones were sequenced, and 12 dominant clones were identified. For conversion to IgG1, the heavy and light chains of Fabs m336, m337, and m338 were amplified and recloned into the pDR12 vector (provided by D. Burton, Scripps Research Institute, La Jolla, CA). Both Fabs and IgG1s were expressed and purified. Protein purity was estimated as >95% by SDS-PAGE, and protein concentration was measured spectrophotometrically (NanoVue, GE Healthcare).
ELISA.
A 96-well plate (Costar) was coated with MERS-CoV RBD (residues 360 to 601) protein at 50 ng/well in phosphate-buffered saline (PBS) overnight at 4°C. For phage ELISA, phage from each round of panning (polyclonal phage ELISA) or clones randomly picked from the infected TG1 cells (monoclonal phage ELISA) were incubated with immobilized antigen. Bound phage were detected with anti-M13-horseradish peroxidase (HRP) polyclonal Ab (Pharmacia, Piscataway, NJ). For the soluble Fab binding assay, HRP-conjugated mouse anti-FLAG tag Ab (Sigma-Aldrich) was used to detect Fab binding. For the IgG1 binding assay, HRP-conjugated goat anti-human IgG Ab (Sigma-Aldrich) was used for detection. For the competition ELISA, 20 nM Fabs were incubated with serially diluted IgG1s, and the mixtures were added to RBD-coated wells. After washing, bound Fabs were detected by HRP-conjugated mouse anti-FLAG tag Ab (Sigma-Aldrich). For inhibition of MERS-CoV RBD binding to DPP4, the plate was coated with recombinant soluble DPP4. The MERS-CoV RBD (residues 377 to 588)-Fc fusion protein was incubated with serially diluted m336, m337, or m338, and the mixtures were added to DPP4-coated wells. After washing, bound RBD-Fc fusion protein was detected by HRP-conjugated goat anti-human IgG Ab (Sigma-Aldrich).
Surface plasmon resonance.
Binding of Fabs and IgG1s to the RBD was assayed using a Biacore X100 instrument. Purified RBD was diluted in 10 mM sodium acetate buffer, pH 5.0, and immobilized on a CM5 sensor chip with an amine coupling kit. The reference flow cell was treated with the amine coupling reagent without exposure to the RBD. The running buffer was HBS-EP (10 mM HEPES, pH 7.4, 150 mM NaCl, 3 mM EDTA, 0.05% surfactant P20). The chip was regenerated with 10 mM glycine, pH 2.5, and 1 M NaCl. The sensorgram was analyzed with BiaEvaluation software, and data were fitted to a 1:1 binding model.
MERS-CoV pseudovirus neutralization assay.
MERS pseudovirus bearing MERS-CoV S protein and a defective HIV-1 genome that expresses luciferase as reporter was prepared by cotransfecting 293T cells with the plasmid pNL4-3.luc.RE (encoding Env-defective, luciferase-expressing HIV-1) and pcDNA3.1-MERS-CoV-S plasmid. To detect the inhibitory activity of MAbs on infection by MERS pseudovirus, DPP4-expressing Huh-7 cells (104/well in 96-well plates) were infected with MERS-CoV pseudovirus in the presence or absence of RBD-specific MAbs m336, m337, and m338 at the indicated concentrations. The culture was refed with fresh medium 12 h postinfection and incubated for an additional 72 h. Cells were washed with PBS and lysed using lysis reagent included in a luciferase kit (Promega). Aliquots of cell lysates were transferred to 96-well flat-bottom luminometer plates (Costar), followed by addition of luciferase substrate (Promega). Relative light units were determined immediately using the Ultra 384 luminometer (Tecan, USA).
MERS-CoV neutralization assay.
A clinical isolate of MERS-CoV was kindly provided by R. Fouchier, A. Zaki, and colleagues. The isolate was cultured in Vero cells with Dulbecco's modified Eagle medium (DMEM) at 37°C. Two or 3 days after virus inoculation, culture supernatants were collected. Serial 2-fold-diluted MAbs m336, m337, and m338 were prepared in duplicate in a 96-well tissue culture plate in MEM supplemented with 1% fetal calf serum (FCS). An equal volume of MERS-CoV working stock containing 200 50% tissue culture infective doses (TCID50) was added, and the antibody-virus mixture was incubated for 2 h at 37°C. Antibody-virus mixtures were added into a 96-well microtiter plate (Costar) containing an equal volume of confluent Vero cells. After incubation at 37°C for 3 days, the plate was observed for cytopathic effect.
Molecular modeling and docking.
The prediction of immunoglobulin structures (PIGS) server (22) was used for automated construction of the three-dimensional (3D) structures of Fv regions of MAbs m336, m337, and m338, using a canonical structure-based approach for complementarity-determining regions (CDRs) L1 to L3, H1, and H2 and grafting of the heavy chain CDR3s (HCDR3s) from known structures. Different criteria for light/heavy chain template selection such as “Same Antibody” and “Best H and L chains” were explored. The conformations of side chains were maintained as long as those residues were conserved between the target and the template, while side chains from other residues were modeled using SCWRL 3.0 as implemented. Finally, the 3D models of MAbs from Fv m336, m337, and m338 were selected without any steric clashes at the VH/VL interface. The ZDOCK 3.0.2 program (23) was employed to dock the MAbs onto the MERS-CoV RBD for which a crystal structure is available. The docking output was filtered by selecting at least one potential contacting residue on the RBD (for example, D539 in RBD) and a residue from the center of the antigen-combining site of MAbs, an Arg residue in the torso region of HCDR3 of m336. The top 10 MERS-CoV RBD docked complexes among the predictions were visually scrutinized. Docked models with the most compatibility to general aspects of antibody-antigen structural features such as hydrogen bonds, salt bridges, and other interactions at the interface without any highly unusual features or clashes were selected.
RESULTS
Selection of high-affinity Fabs to the MERS-CoV RBD from a very large naive human phage-displayed antibody library.
To select high-affinity antibodies without additional maturation, we constructed a very large phage-displayed antibody Fab library (containing ∼1011 antibodies) made by using B cells from the blood of 40 healthy donors. Preliminary characterization of this library indicated very high diversity, demonstrated also by identification of high-affinity binders toward various viral and cancer-related antigens. This library was panned against recombinant MERS-CoV RBD developed in our laboratory (Fig. 1). Panning resulted in significant enrichment after 3 or 4 rounds. Screening against the same protein led to the identification of a panel of 12 Fabs that bound with various affinities to the RBD as demonstrated by ELISA and Biacore (Fig. 1 and 2). Three Fabs designated m336, m337, and m338 had the lowest concentrations of 50% binding (EC50s). Interestingly, we found that all three MAbs emerged from the common germ line VH gene 1-69 (Table 1).
FIG 1.

(A) Schematic structure of MERS-CoV S subunit and RBD(360-601)-Avi tag. SP, signal peptide. RBD(360-601)-Avi tag was constructed by fusing MERS-CoV residues 360 to 601 of S1 with Avi tag. (B) Polyclonal phage ELISA showing the binding of the first to fourth rounds (Rd 1 to Rd 4) of phage to RBD. Bound phage were detected with anti-M13-HRP polyclonal Ab. (C) Binding of Fabs m331 to m342 to RBD. HRP-conjugated mouse anti-FLAG tag Ab was used to detect Fab binding.
FIG 2.

Binding of Fabs m336, m337, and m338 to MERS-CoV RBD measured by BIAcore. RBD was immobilized on a CM5 chip. The analytes consisted of serial dilution of proteins between 0.5 μM and 0.8 nM. The chip was regenerated with 10 mM glycine (pH 2.5)–1 M NaCl.
TABLE 1.
Gene usage and binding kinetics features of MERS-CoV RBD-specific Fabs and IgG1s
| Feature (unit) | m336 | m337 | m338 |
|---|---|---|---|
| Gene families | |||
| VH | 1-69*06 | 1-69*06, 08 or 14 | 1-69*09 |
| DH | 2-2*03 | 3-22*01 | 3-22*01 |
| JH | J3*02 | J3*02 | J3*02 |
| VLa | 1-17*01 | 2-28*01, 2D-28*01 | 3-11*01 |
| JL | 4*01 | 4*01 | 1*01 |
| Binding kinetics for Fabs | |||
| kon (M · s−1) | 4.50 × 106 | 4.49 × 104 | 3.68 × 104 |
| koff (s−1) | 1.92 × 10−2 | 4.19 × 10−4 | 5.54 × 10−4 |
| Kd (M) | 4.27 × 10−9 | 9.34 × 10−9 | 1.51 × 10−8 |
| Binding kinetics for IgG1s | |||
| kon (M−1 · s−1) | 1.66 × 106 | 1.87 × 105 | 3.55 × 105 |
| koff (s−1) | 1.65 × 10−4 | 1.54 × 10−4 | 1.98 × 10−4 |
| Kd (M) | 9.94 × 10−11 | 8.23 × 10−10 | 5.59 × 10−10 |
All three are kappa light chains.
Generation of full-size antibodies in IgG1 format and their binding to the RBD.
Fabs m336, m337, and m338 were selected for further characterization and converted to an IgG1 format. The MAbs bound with high (pM) avidity (dissociation constant [Kd], 9.94 × 10−11 to 8.23 × 10−10 M) to RBD as measured by Biacore (Table 1), but the negative-control IgG1, m610.27, which has the same constant domains as m336, m337, and m338, did not (data not shown). Their high affinity/avidity and specificity suggest potential use as research reagents for exploring mechanisms of viral entry and for diagnosis of MERS-CoV infections.
Neutralization of MERS-CoV infection by RBD-specific MAbs.
The neutralization activities of IgG1s m336, m337, and m338 against MERS-CoV were first evaluated by using a pseudovirus system. Single-round HIV luciferase reporter viruses pseudotyped with MERS-CoV S protein were tested for neutralization sensitivity against increasing concentrations of MAbs. As shown in Fig. 3A, all three RBD-specific MAbs exhibited exceptionally potent neutralization activity, with 50% inhibitory concentration (IC50) ranging from 0.005 to 0.017 μg/ml. Notably, the most potent MAb, m336, inhibited >90% MERS-CoV pseudovirus infection at a concentration of 0.039 μg/ml, while the negative-control IgG1, m610.27, was unable to inhibit the infection (data not shown).
FIG 3.

Potent in vitro neutralization of MERS-CoV by RBD-specific MAbs. (A) Neutralization of viruses pseudotyped with the MERS-CoV S glycoprotein. Pseudotyped virus was incubated with IgG1s before infection of DPP4-expressing Huh-7 cells. Luciferase activities were measured, and percent neutralization was calculated. (B) Neutralization of live virus. MERS-CoV was incubated with IgG1s before infection of Vero cells. Cytopathic effects were measured, and percent neutralization was calculated.
We next tested the neutralization activity of the MAbs with live MERS-CoV virus (Fig. 3B). Similarly, m336 showed the most potent neutralization activity, with IC95 of 1 μg/ml and IC50 of 0.07 μg/ml. To our knowledge, these are the first fully human MAbs known to neutralize both pseudotyped and live MERS-CoV with such high potency.
RBD-specific MAbs competed with the MERS-CoV receptor DPP4 for binding to the virus.
We used several S1 fragments, a soluble DPP4 (sDPP4), alanine mutagenesis, and molecular modeling to define the epitopes of the antibodies and elucidate the mechanism of neutralization.
MERS-CoV S fragments S18 to S725 (S18-725), S18-353, S358-588, S377-588, S377-662, and S726-1295 fused with the Fc fragment of IgG1 were generated (Fig. 4A). As shown in Fig. 4B to D, m336, m337, and m338 bound potently to fragments containing the RBD (S18-725-Fc, S358-588-Fc, S377-588-Fc, and S377-662-Fc) but not to fragments lacking the RBD (S726-1295-Fc and S18-353-Fc). These data confirm that the binding sites of these MAbs are within the MERS-CoV RBD and, specifically, in the region of residues 377 to 588.
FIG 4.

Binding of Fabs to different MERS-CoV S segments. (A) Schematic structure of MERS-CoV S subunits S18-725, S18-353, S358-588, S377-588, S377-662, and S726-1295 fused with the Fc fragment of IgG1. m336 (B), m337 (C), and m338 (D) bound to S18-725-Fc, S358-588-Fc, S377-588-Fc, and S377-662-Fc but not to S726-1295-Fc or S18-353-Fc.
We next evaluated whether MAbs m336, m337, and m338 were capable of inhibiting the binding of MERS-CoV RBD to the receptor DPP4. We first confirmed that the three MAbs competed with each other for binding to the MERS-CoV S1 glycoprotein (Fig. 5A). Furthermore, as shown in Fig. 5B, all the IgG1s potently inhibited the binding of RBD to the soluble DPP4 receptor. Consistent with the neutralization results, m336 IgG1 was slightly more potent than m337 and m338 in blocking the binding of RBD to the receptor. The IC50s of m336, m337, and m338 were 0.034, 0.044, and 0.041 μg/ml, respectively. Taken together, these results suggest that the MAbs neutralize the virus by competing with the receptor for binding to the RBD and have overlapping epitopes.
FIG 5.

MAbs competed with the MERS-CoV receptor DPP4 for binding to RBD. (A) m336, m337, and m338 competed with each other for binding to the MERS-CoV RBD. Fabs were incubated with serially diluted IgG1s, and the mixtures were added to RBD-coated wells. After washing, bound Fabs were detected by HRP-conjugated mouse anti-FLAG tag Ab. (B) The MERS-CoV RBD-Fc fusion protein was incubated with serially diluted IgG1s, and the mixtures were added to DPP4-coated wells. After washing, bound RBD-Fc fusion protein was detected by HRP-conjugated goat anti-human IgG Ab.
To further localize the MAb epitopes, a panel of RBD alanine-scanning mutants was developed, guided by structural information available for the RBD. Binding of MAbs to the mutants were measured by ELISA (Fig. 6). Binding of m336 to RBD with mutated residues L506A, D510A, E536A, D539A, W553A, and V555A was considerably reduced, as were binding of m337 to mutants L506A, D510A, E553A, and V555A and binding of m338 to mutants D510A and W553A. These results indicate that the three RBD-specific MAbs possess overlapping but distinct epitopes. In addition, the most potent MAb, m336, uniquely lost binding to the E536/D539 mutant, indicating that these RBD residues are crucial for interactions with m336 and could be part of a promising template for the development of vaccine immunogens. In contrast, residues D510 and W553 appear to be important for RBD binding to all three MAbs.
FIG 6.

Binding of Fabs m336 (A), m337 (B), and m338 (C) to different MERS-CoV RBD mutants.
Molecular modeling of RBD-specific MAbs and docking with the MERS-CoV RBD.
We used the mutagenesis data and the RBD crystal structure (24, 25) to build docking models (Fig. 7). Both the Fv regions of light and heavy chains of MAbs m336, m337, and m338 were modeled using the best matches with highest sequence identities of known templates from different structures. All the CDRs except HCDR3 were modeled based on canonical conformations defined for those particular canonical structure classes. Although there are no such canonical structures for HCDR3, HCDR3 of RBD-specific MAbs were grafted from structurally known antibodies with the same HCDR3 lengths. This combined approach resulted in the 3D models of RBD-specific MAbs as shown in Fig. 7A to C. In the case of m336, the HCDR3 template (PDB code 3EYF) also had centrally located cysteine residues that formed a disulfide bond which could be used to stabilize the conformation of HCDR3 with a length of 18 amino acids in m336. The other two MAbs, m337 and m338, had multiple tyrosine residues within the HCDR3s that might be of structural significance as well as active antigen recognition sites. For docking simulations, at least one of the HCDR3 residues, which was well exposed to solvent and located at the middle of the antigen combining site, was considered a contacting residue in m336. Similarly, one of the most prominent residues identified from the site-directed mutagenesis experiment, which was also located at the RBD surface, was considered a contacting residue in the MERS-CoV RBD. The similar docking constraints for m337 and m338 helped in selecting the docked complexes that might predict potential binding residues at the interface of MERS-CoV RBD-MAbs complexes (Table 2). The known crystal structure of MERS-CoV RBD bound to DPP4 (24, 25) was superimposed onto each of the three MAbs by aligning the RDB region as shown in Fig. 7D. These results indicated a possible dominant role of the heavy chain in the MAb paratopes and putative epitope-forming RBD residues (Table 2), showing the overlap between the MAb epitopes and the receptor binding site (Fig. 7D) and molecular details of the MAb-RBD interactions that may have implications for design of vaccine immunogens and small-molecule drugs.
FIG 7.
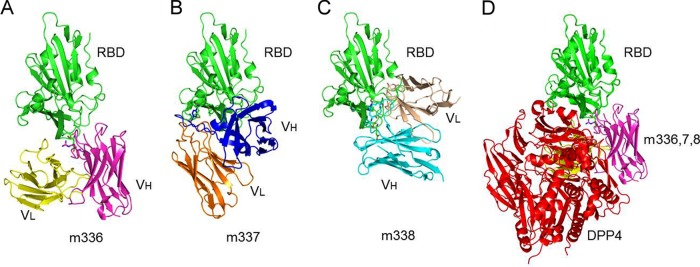
Docked complexes of MERS-CoV RBD with MAbs m336 (A), m337 (B), and m338 (C). (D) Superposition of the docked complexes of RBD-m336, -m337, and -m338 and crystal structure of the RBD-DPP4 complex.
TABLE 2.
Putative binding residues predicted from the docked complexes of MERS-CoV RBD and m336, m337, and m338a
| MERS-CoV RBD-m336 |
MERS-CoV RBD-m337 |
MERS-CoV RBD-m338 |
|||
|---|---|---|---|---|---|
| RBD residue | MAb m336 residue | RBD residue | MAb m337 residue | RBD residue | MAb m338 residue |
| Lys493 | Thr28 | Phe467 | Ser31H | Leu495 | Ser31 |
| Leu495H | Ser30 | Asn501 | Tyr32 | Ile529 | Arg50 |
| Lys496 | Tyr32 | Lys502H | Ile52H | Ser532H | Ile52 |
| Y499 | Tyr101 | Leu506 | Phe55 | Pro531 | Leu55 |
| Asn501 | Ser104 | Asp510H | Thr57H | Thr533 | Ile57 |
| Thr533 | Thr105 | Arg511H | Asn59 | Trp535H | Asn59 |
| Trp535 | Arg109HS | Glu513S | Gln65H | Glu536H | Asp102 |
| Glu536HS | Ala111H | Pro531 | Leu100 | Asp539 | Ser104H |
| ASP537 | Asp113H | Trp535 | Asp105 | Tyr540 | Tyr106 |
| Asp539HS | Ile114 | Glu536HS | Tyr110 | Tyr541 | Tyr107 |
| Tyr540 | Asp539H | Arg542H | Arg108 | ||
| Arg542 | Tyr49 | Tyr540H | Gln27 | Lys543 | |
| Val561 | Gln55 | Arg542HS | His31S | Gln544H | Gln27 |
| Met563 | Ser56 | Lys543 | Leu97 | Trp553 | His31S |
| Glu565 | Gln544H | Gln98 | Leu97 | ||
| Trp553 | Thr99 | Gln98 | |||
| Thr99 | |||||
H and HS, possible hydrogen bonds and hydrogen bonds/salt bridges, respectively, across the molecular interface; in m336, m337, and m338 columns, lightface indicates VL and boldface indicates VH.
DISCUSSION
The major results of this study are the identification and characterization of novel human MAbs against the recently emerged MERS-CoV that bind to the MERS-CoV RBD with very high affinity (pM) and inhibit infection with exceptional potency. We attempted to test the antibody inhibitory activity in mice, but the level of virus was very low likely due to the inefficient or nonexistent virus entry mediated by the mouse DPP4 (26). Therefore, a better model should be used to evaluate the antibody potency in vivo. Interestingly, the Ig heavy chain variable region (IGHV) genes of m336,m337, and m338 originated from IGHV1-69. It is noted that the IGHV1-69 gene is also preferentially utilized by other antiviral antibodies, including HIV-1 (27), influenza virus (28), and hepatitis C virus (29).
We found that that the three MAbs compete with each other and with the receptor DPP4 for binding to the S glycoprotein, suggesting a competitive mechanism of virus neutralization. Several other MAbs against emerging viruses, including SARS-CoV and henipaviruses, which exhibited high potency in vitro and in animal models of infection, have been previously identified (18–20) and used in humans exposed to Hendra virus (30). Because they competed with the receptors and some bound to the receptor binding site (RBS), they were highly cross-reactive. For example, m102.4 inhibited both Hendra and Nipah viruses, which use the same receptor for entry into cells. Therefore, one can expect that the newly identified antibodies will be effective against many, if not all, MERS-CoV isolates. We analyzed the sequences of a total of 11 MERS-CoV strains and found that overall their sequences are highly conserved. Only 5 residues of the MERS-CoV S1 glycoprotein (725 residues in length) and only 1 residue in the RBD region contained any variation at all. In addition, one could expect that the emergence of resistant viruses, if any, will exert a toll on the virus fitness because the mutant virus may have lower affinity for the receptor. However, further experiments to test m336 against different isolates as well as against emergence of resistant mutants are needed to confirm this possibility.
These newly identified MAbs are promising candidate therapeutics against the MERS-CoV because of their exceptionally high neutralization activity, which could accelerate their further development for prophylaxis and therapy of MERS-CoV infection in humans. We also expect their uses as a reagent to facilitate the development of highly immunogenic vaccines against MERS-CoV. We have found that significant neutralizing antibody responses could be induced by immunizing BALB/c mice with purified MERS-CoV RBD protein (31). Taken together, these results suggest that RBD, especially the epitopes that are targeted by these exceptionally potent RBD-specific MAbs, has great potential to be developed as a highly effective MERS-CoV vaccine. Additionally, the rapid identification (within several weeks) of three potent MAbs described in this work suggests a possibility to use the large nonimmune antibody library and related methodology for a quick response to the public threat resulting from emerging SARS-like or MERS-like coronaviruses, which are highly likely to continue to emerge and pose a serious threat to human health in the near future. MERS-CoV continues to cause major illness and has even killed a number of people in close contact with patients, as has been reported (2, 32). These MAbs may help treat patients in similar situations and can be a life-saving intervention for those who have been infected or are at risk of MERS-CoV infections.
ACKNOWLEDGMENTS
We thank Fang Li from the University of Minnesota for useful discussions.
This work was supported by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research, and by Federal funds from the NIH, National Cancer Institute, under contract no. N01-CO-12400 and HHSN261200800001E.
Footnotes
Published ahead of print 30 April 2014
REFERENCES
- 1.Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. 2012. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 367:1814–1820. 10.1056/NEJMoa1211721 [DOI] [PubMed] [Google Scholar]
- 2.Assiri A, McGeer A, Perl TM, Price CS, Al Rabeeah AA, Cummings DA, Alabdullatif ZN, Assad M, Almulhim A, Makhdoom H, Madani H, Alhakeem R, Al-Tawfiq JA, Cotten M, Watson SJ, Kellam P, Zumla AI, Memish ZA. 2013. Hospital outbreak of Middle East respiratory syndrome coronavirus. N. Engl. J. Med. 369:407–416. 10.1056/NEJMoa1306742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cotten M, Watson SJ, Kellam P, Al-Rabeeah AA, Makhdoom HQ, Assiri A, Al-Tawfiq JA, Alhakeem RF, Madani H, AlRabiah FA, Al Hajjar S, Al-nassir , Albarrak WNA, Flemban H, Balkhy HH, Alsubaie S, Palser AL, Gall A, Bashford-Rogers R, Rambaut A, Zumla AI, Memish ZA. 2013. Transmission and evolution of the Middle East respiratory syndrome coronavirus in Saudi Arabia: a descriptive genomic study. Lancet 382:1993–2002. 10.1016/S0140-6736(13)61887-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ithete NL, Stoffberg S, Corman VM, Cottontail VM, Richards LR, Schoeman MC, Drosten C, Drexler JF, Preiser W. 2013. Close relative of human Middle East respiratory syndrome coronavirus in bat, South Africa. Emerg. Infect. Dis. 19:1697–1699. 10.3201/eid1910.130946 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Haagmans BL, Al Dhahiry SH, Reusken CB, Raj VS, Galiano M, Myers R, Godeke GJ, Jonges M, Farag E, Diab A, Ghobashy H, Alhajri F, Al-Thani M, Al-Marri SA, Al Romaihi HE, Al Khal A, Bermingham A, Osterhaus AD, Alhajri MM, Koopmans MP. 2014. Middle East respiratory syndrome coronavirus in dromedary camels: an outbreak investigation. Lancet Infect. Dis. 14:140–145. 10.1016/S1473-3099(13)70690-X [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Graham RL, Donaldson EF, Baric RS. 2013. A decade after SARS: strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 11:836–848. 10.1038/nrmicro3143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Falzarano D, de Wit E, Rasmussen AL, Feldmann F, Okumura A, Scott DP, Brining D, Bushmaker T, Martellaro C, Baseler L, Benecke AG, Katze MG, Munster VJ, Feldmann H. 2013. Treatment with interferon-alpha2b and ribavirin improves outcome in MERS-CoV-infected rhesus macaques. Nat. Med. 19:1313–1317. 10.1038/nm.3362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dimitrov DS. 2012. Therapeutic proteins. Methods Mol. Biol. 899:1–26. 10.1007/978-1-61779-921-1_1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schrama D, Reisfeld RA, Becker JC. 2006. Antibody targeted drugs as cancer therapeutics. Nat. Rev. Drug Discov. 5:147–159. 10.1038/nrd1957 [DOI] [PubMed] [Google Scholar]
- 10.Casadevall A, Dadachova E, Pirofski L. 2004. Passive antibody therapy for infectious diseases. Nat. Rev. Microbiol. 2:695–703. 10.1038/nrmicro974 [DOI] [PubMed] [Google Scholar]
- 11.Carter PJ. 2006. Potent antibody therapeutics by design. Nat. Rev. Immunol. 6:343–357. 10.1038/nri1837 [DOI] [PubMed] [Google Scholar]
- 12.Scott LJ, Lamb HM. 1999. Palivizumab. Drugs 58:305–313. 10.2165/00003495-199958020-00009 [DOI] [PubMed] [Google Scholar]
- 13.Prabakaran P, Zhu Z, Xiao X, Biragyn A, Dimitrov AS, Broder CC, Dimitrov DS. 2009. Potent human monoclonal antibodies against SARS CoV, Nipah and Hendra viruses. Expert Opin. Biol. Ther. 9:355–368. 10.1517/14712590902763755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Marasco WA, Sui J. 2007. The growth and potential of human antiviral monoclonal antibody therapeutics. Nat. Biotechnol. 25:1421–1434. 10.1038/nbt1363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kwong PD, Mascola JR. 2012. Human antibodies that neutralize HIV-1: identification, structures, and B cell ontogenies. Immunity 37:412–425. 10.1016/j.immuni.2012.08.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhu Z, Dimitrov AS, Bossart KN, Crameri G, Bishop KA, Choudhry V, Mungall BA, Feng YR, Choudhary A, Zhang MY, Feng Y, Wang LF, Xiao X, Eaton BT, Broder CC, Dimitrov DS. 2006. Potent neutralization of Hendra and Nipah viruses by human monoclonal antibodies. J. Virol. 80:891–899. 10.1128/JVI.80.2.891-899.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bossart KN, Tachedjian M, McEachern JA, Crameri G, Zhu Z, Dimitrov DS, Broder CC, Wang LF. 2008. Functional studies of host-specific ephrin-B ligands as Henipavirus receptors. Virology 372:357–371. 10.1016/j.virol.2007.11.011 [DOI] [PubMed] [Google Scholar]
- 18.Bossart KN, Zhu Z, Middleton D, Klippel J, Crameri G, Bingham J, McEachern JA, Green D, Hancock TJ, Chan YP, Hickey AC, Dimitrov DS, Wang LF, Broder CC. 2009. A neutralizing human monoclonal antibody protects against lethal disease in a new ferret model of acute nipah virus infection. PLoS Pathog. 5:e1000642. 10.1371/journal.ppat.1000642 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bossart KN, Geisbert TW, Feldmann H, Zhu Z, Feldmann F, Geisbert JB, Yan L, Feng YR, Brining D, Scott D, Wang Y, Dimitrov AS, Callison J, Chan YP, Hickey AC, Dimitrov DS, Broder CC, Rockx B. 2011. A neutralizing human monoclonal antibody protects African green monkeys from Hendra virus challenge. Sci. Transl. Med. 3:105ra103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhu ZY, Chakraborti S, He Y, Roberts A, Sheahan T, Xiao XD, Hensley LE, Prabakaran P, Rockx B, Sidorov IA, Corti D, Vogel L, Feng Y, Kim JO, Wang LF, Baric R, Lanzavecchia A, Curtis KM, Nabel GJ, Subbarao K, Jiang S, Dimitrov DS. 2007. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc. Natl. Acad. Sci. U. S. A. 104:12123–12128. 10.1073/pnas.0701000104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Du L, Kou Z, Ma C, Tao X, Wang L, Zhao G, Chen Y, Yu F, Tseng CT, Zhou Y, Jiang S. 2013. A truncated receptor-binding domain of MERS-CoV spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: implication for developing therapeutics and vaccines. PLoS One 8:e81587. 10.1371/journal.pone.0081587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Marcatili P, Rosi A, Tramontano A. 2008. PIGS: automatic prediction of antibody structures. Bioinformatics 24:1953–1954. 10.1093/bioinformatics/btn341 [DOI] [PubMed] [Google Scholar]
- 23.Chen R, Li L, Weng Z. 2003. ZDOCK: an initial-stage protein-docking algorithm. Proteins 52:80–87. 10.1002/prot.10389 [DOI] [PubMed] [Google Scholar]
- 24.Lu G, Hu Y, Wang Q, Qi J, Gao F, Li Y, Zhang Y, Zhang W, Yuan Y, Bao J, Zhang B, Shi Y, Yan J, Gao GF. 2013. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 500:227–231. 10.1038/nature12328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang N, Shi X, Jiang L, Zhang S, Wang D, Tong P, Guo D, Fu L, Cui Y, Liu X, Arledge KC, Chen YH, Zhang L, Wang X. 2013. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 23:986–993. 10.1038/cr.2013.92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Devitt E. 2013. Lack of small animal model hinders MERS coronavirus research. Nat. Med. 19:952. 10.1038/nm0813-952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Prabakaran P, Zhu Z, Chen W, Gong R, Feng Y, Streaker E, Dimitrov DS. 2012. Origin, diversity, and maturation of human antiviral antibodies analyzed by high-throughput sequencing. Front. Microbiol. 3:277. 10.3389/fmicb.2012.00277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ohshima N, Iba Y, Kubota-Koketsu R, Asano Y, Okuno Y, Kurosawa Y. 2011. Naturally occurring antibodies in humans can neutralize a variety of influenza virus strains, including H3, H1, H2, and H5. J. Virol. 85:11048–11057. 10.1128/JVI.05397-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chan CH, Hadlock KG, Foung SK, Levy S. 2001. V(H)1–69 gene is preferentially used by hepatitis C virus-associated B cell lymphomas and by normal B cells responding to the E2 viral antigen. Blood 97:1023–1026. 10.1182/blood.V97.4.1023 [DOI] [PubMed] [Google Scholar]
- 30.Zhu Z, Prabakaran P, Chen W, Broder CC, Gong R, Dimitrov DS. 2013. Human monoclonal antibodies as candidate therapeutics against emerging viruses and HIV-1. Virol. Sin. 28:71–80. 10.1007/s12250-013-3313-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Du L, Zhao G, Kou Z, Ma C, Sun S, Poon VK, Lu L, Wang L, Debnath AK, Zheng BJ, Zhou Y, Jiang S. 2013. Identification of a receptor-binding domain in the S protein of the novel human coronavirus Middle East respiratory syndrome coronavirus as an essential target for vaccine development. J. Virol. 87:9939–9942. 10.1128/JVI.01048-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Memish ZA, Zumla AI, Al-Hakeem RF, Al-Rabeeah AA, Stephens GM. 2013. Family cluster of Middle East respiratory syndrome coronavirus infections. N. Engl. J. Med. 368:2487–2494. 10.1056/NEJMoa1303729 [DOI] [PubMed] [Google Scholar]