

Abstract
A thorough understanding of nerve regeneration in Caenorhabditis elegans requires performing femtosecond laser nanoaxotomy while minimally affecting the worm. We present a microfluidic device that fulfills such criteria and can easily be automated to enable high-throughput genetic and pharmacological screenings. Using the ‘nanoaxotomy’ chip, we discovered that axonal regeneration occurs much faster than previously described and surprisingly the distal fragment of the severed axon regrows in the absence of anesthetics.
The understanding of the biological mechanisms of nerve regeneration and degeneration after injury holds the key to developing novel therapies for human neurodegenerative diseases. These processes can be studied in model organisms by severing axons in a controlled manner, and then observing their regrowth and functional recovery. The ideal predisposition of the nematode Caenorhabditis elegans for such studies recently became accessible by the demonstration of precise nanoaxotomy using ultrafast laser pulses1. However, the side effects that the chemicals used to immobilize the worms for laser nanoaxotomy might have on nerve regeneration are difficult to evaluate, unless nanosurgery can be performed in vivo without anesthetics. The environment in which both surgery and monitoring are performed should have a minimal impact on the studied organism and its biological processes. To achieve this goal, we designed a microfluidic device that allows us to sever axons in C. elegans using ultrashort laser pulses with the same high precision as we demonstrated previously1,2 while monitoring the subsequent axonal regeneration activity.
Several microfluidic devices and microelectromechanical systems (MEMS) have recently been developed for C. elegans including, Petri dish-based microfluidics3, microfluidic traps4,5,6, the “CD-worm”7, a shadow imaging platform8, microfluidic maze structures6,9, a cantilever force MEMS sensor10 and a platform to capture and sort worms11. However no demonstration of nanosurgery on-a-chip has been reported so far.
The integrated microfluidic device we designed presents several unique features that are critical for the success of in vivo nerve regeneration studies: the worms are held directly against the glass cover for ideal focusing and precise nanosurgery; the trap is adjustable to the size of the worms allowing immobilization of worms at various developmental stages (L4 to adult size); and the system integrates feeding modules and thus allows long term follow-up studies of the axotomized worms as well as their sorting and screening.
The high-throughput microfluidic system integrates two separate modules (Fig. 1a), a trapping module for nanosurgery and time–lapse imaging (Fig. 1c–e) and a feeding module for recovery of the operated worms (Fig. 1b). Follow-up imaging of injured axons and their regrowth is performed using the same trapping module. Depending on the outcome of the imaging session, the worms can either be flushed out through the inlet or sorted into different feeding chambers according to their axonal recovery progress. If necessary, the feeding chambers can be connected to a supply of fresh liquid growth medium, allowing observations for an extended period of time. The whole cycle of loading, trapping, and operating a worm takes approximately 1 minute whereas it takes 10 minutes per worm using the previous technique of nanoaxotomy on agar pads1.
Figure 1.

The nanoaxotomy lab-on-a-chip. (a) Overview of the chip with the trap system (yellow rectangle) and three recovery chambers (blue rectangle). (b) Magnified view of recovery chambers. Valves 5, 6 and 7 (yellow rectangles) work in pairs and regulate the loading of a worm to either chamber accordingly. (c) Magnified view of the trapping system. Valves 1 to 4 (yellow rectangles) respectively control inlet regulation, fine positioning of the worm (2 & 3) and gating to the recovery chambers. (d) Conceptual 3D sectional renderings of the bilayer trap channels without and with an immobilized worm. (e) Two-photon images of cross-sectional profiles of the microchannel in the trap area for increasing air pressures from 0 to 35, 70, 105, 140 and 175 kPa. (f) Cross-sectional two-photon images of a trapped worm at 105 and 140 kPa. Scale bars are 2 mm in (a) and (b), 1 mm in (c), and 50 μm in (e) and (f).
We adopted a two-layer approach12 that incorporates a thin membrane (~40 μm) between the two layers of microfluidic channels (Supplementary Methods). The worms are loaded in a 50 μm deep and 110 μm wide microfluidic channel in the bottom layer of the chip (Fig 1c). Pressurization of the top layer results in the deflection of the membrane that quickly immobilizes the freely moving animal (Supplementary Video 1). After the axons of the immobilized animal are severed, the pressure is released and the animal is transferred to one of the feeding chambers.
Membrane deflection depends on the applied pressure in the air channel as well as the dimensions of the bottom channel. Using two-photon imaging (Supplementary Methods), we mapped the cross-sectional profile of the trapping channel and studied the deflection of the membrane at different pressures (Fig. 1e). As pressure increases in the top layer, the membrane begins deflecting downward, immobilizing the nematode by pressing it into one of the sides of the channel (Fig. 1d,f). Immobilization of L4 and young adults was achieved using a minimal pressure of 110 kPa air. We performed a survival test on 20 worms, trapped with a pressure of 110 kPa for 60 minutes. We monitored them for the following three days and did not observe any morphological and behavioral defects or premature death (Table 1, experimental set 5).
Table 1.
Summary of statistical observations. We performed nanoaxotomy and monitored the axonal recovery (regrowth and reconnection) of groups of 20 worms grown either on agar or in liquid growth medium (CeRH) and either paralyzed by a 0.2 μM solution of levamisole and operated on agar pads or trapped in the new nanoaxotomy lab-on-a-chip device.
| Set # | Growth medium | Surgery medium | Surgery on agar pad, paralyzed | Trapping on-a-chip | Surgery on-a-chip | Survival Rate(worms), 24 hrs | Axonal recovery time | Recovery Rate(axons), 24 hrs | Survival Rate (worms), 72 hrs |
|---|---|---|---|---|---|---|---|---|---|
| 1 | agar | agar | + | 20/20 (100%) | 6 –12 hrs | 18/40 (45%) | 18/20 (90 %) | ||
| 2 | agar | M9 | + | + | 20/20 (100%) | 60 – 90 min | 24/40 (60%) | 20/20 (100%) | |
| 3 | CeRH | agar | + | 20/20 (100%) | 6 – 12 hrs | 21/40 (53%) | 20/20 (100%) | ||
| 4 | CeRH | CeRH | + | + | 20/20 (100%) | 60 – 90 min | 26/40 (65%) | 20/20 (100%) | |
| 5 | CeRH | no | + | 20/20 (100%) | N/A | N/A | 20/20 (100%) |
The cross-sectional view of the channel with a trapped animal (Fig. 1f) shows that the side of the nematode lying against the cover glass is flattened. This flattening is optimal for both imaging and surgery since most of the length of the target axon will be in focus. This design provides an optical path from the objective lens to the axon inside the animal that is similar to the previous nanoaxotomy method on agar pads1,2.
Nanosurgery of neuronal processes was performed by a train of 200 highly focused 220-fs long laser pulses of 7.2 nJ at 1 kHz repetition rate and 780 nm wavelength (Supplementary Methods and Supplementary Video 2). Axotomies were performed at 1/3 of the process length from cell body to axon branching. For time lapse imaging, fluorescence images were taken at approximately ten minute intervals (Fig. 2). We observed that the regrowth was not continuous but rather it occurred in bursts of elongation and retraction of filopodia that could be as fast as 2 μm in 10 seconds. Axons reconnected to their distal end within approximately 70 minutes after surgery.
Figure 2.
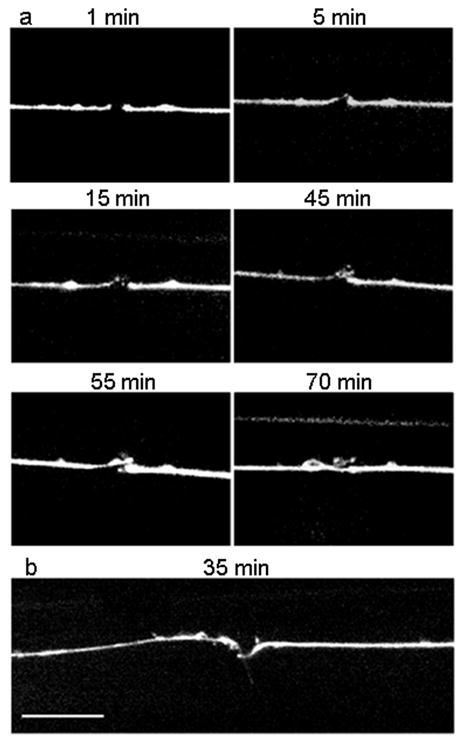
Time-lapse imaging of axonal recovery on-a-chip. (a) Fluorescence images of an ALM neuron at 1, 5, 15, 45, 55 and 70 minutes after the axotomy, respectively. Distal ends are on the left side of the pictures, proximal ones on the right. After 5 minutes, the distal end displays a growth cone that is visible above the proximal stump. At 15 minutes, the growth cone branches off into two. At 45 minutes, a third branch sprouts close to the distal stump. The other two growth cones are out of focus. At 55 minutes, the third branch recesses and the first two branches develop into a broad growth cone. The proximal end starts regrowing. At 70 minutes, the proximal end regrew and reconnected to the distal end a bit further past the distal stump. (b) Fluorescence image of axonal recovery in a different axon 35 minutes after axotomy. The distal growth was misguided and developed sideways (pointing downward on the picture). The proximal end is regrowing towards the distal part. Scale bar is 10 μm.
In order to investigate whether the absence of anesthetics (phenoxypropanol or levamisole) during nanosurgery would increase the probability of successful axonal recovery (regrowth and reconnection), we performed four sets of experiments (Table 1, experimental sets 1–4). We first considered the effect of the growth medium on axonal recovery. Worms grown on agar and in CeRH liquid growth medium both showed similar axonal recovery rates. Considering the growth media as the variable, the Fisher’s exact test (Supplementary Methods) yields probabilities of P13 = 0.284 for worms axotomized on agar pad using levamisole (experimental sets 1 and 3), P24 = 0.328 for worms axotomized in chip using the microfluidics trap (experimental sets 2 and 4), and P13–24 = 0.184 regardless of the immobilization procedure. The change of the growth environment thus seems to have no statistical significance.
To characterize the statistical significance of the absence of anesthetics on nerve regeneration, we next compared the experiments differing on the immobilization method. Considering the immobilization procedure as the variable, the Fisher’s exact test yields the following probabilities: P12 = 0.146, P34 = 0.192, and P12–34 = 0.046 for worms grown on agar, in CeRH liquid medium, and regardless of the growth habitat, respectively. A probability of 0.046 indicates that as expected, anesthetics significantly slow down the nerve regeneration processes and delay it by several hours2.
To our surprise, in addition to regrowth of the proximal fragments, we also observed the distal fragments always sprouting growth cones. The regrowth of the distal fragment was already visible within a few minutes after axotomy, while it takes at least 30 minutes for the proximal fragment to start regrowing. The regrowth velocity of both fragments was the same. Interestingly, the distal ends seemed to lack any kind of guidance and never reconnected to the proximal end. The regrowth of the distal end seemed to stop shortly after the proximal end began its regrowth. Furthermore, once the proximal growth cones reconnected to their distal ends, the growth cones from the distal ends did not recess (Fig. 2b). This result suggests that the microtubules within the distal end are stable and that actin, mitochondria, and proteins necessary for the creation of a growth cone are present or might be transported from the remaining section of the process to its severed end. Because the cell body is missing and membranous vesicles cannot be fabricated, the mobility of the growth cone requires that these vesicles be endocytosed, transported, and exocytosed at the forefront of the growth cone13. The hypodermal syncytium hyp7, enclosing the ALM and PLM neurons, is thought to provide a structural cue for the guidance of the proximal growth cones14.
We also observed that axonal recovery does not depend on the location of the surgery along the process and even immediately distal to the axonal branch (Supplementary Methods). Axonal recovery rates varied between 62% and 73% with no statistically significant difference (P = 0.627). In a recent study15, Wu et al. showed that there was no regeneration of a severed axon beyond the axonal branch (the “distal process” as named by Wu et al.) unless the branch is cut to stimulate regrowth of the distal process. We also performed surgeries on the branch and on both branch and distal process, both in chip and on agar pads. We did not observe any axonal recovery of severed branches in agreement with Wu et al. However, we conclude that regeneration of the distal process occur independent of whether the branch is cut or not.
Behavioral assays are a widespread method to ascertain function of specific neurons. We proved the reliability of the laser surgery by ablating the circumferential axons of the VD and DD motor neurons in the microfluidic device and observed the shrinker phenotype during a behavioral assay. We also tested the behavioral output of worms after severing the ALM and PLM axons at various locations. In general, C. elegans respond with a clear backward movement to a light mechanical stimulus (light touch) applied on the anterior half of their body (ALM mediated) and with a forward acceleration when the light touch is applied on the posterior half of the body (PLM mediated). With axotomies performed on different sections of ALM and PLM processes we have found that the cell body as well as a large part of the process themselves are not essential for a behavioral response to occur. Worms responded well with no change in their behavior when (1) ALM and PLM processes cut on one side only, (2) ALM and PLM processes cut on both sides, and (3) cell bodies of both ALM and PLM pairs ablated. On the other hand, axotomy at the branch clearly impaired the mechanical response (Supplementary Video 3). These results confirm the dual axonal-dendritic nature of the mechanosensory processes and demonstrate the crucial function of the synaptic branch to generate the light touch behavioral output.
The described microfluidic chip possesses many advantages over the immobilization techniques that were previously used in studies of C. elegans, such as anesthesia on agar pads or glue, including: (1) no chemicals other than the liquid growth medium will interfere with the physiological processes of the worms, possibly increasing regeneration success, (2) the adaptive deflection of the membrane allows the immobilization of the worms from L4 to adult size, (3) the worms do not need a recovery period after surgery, permitting immediate behavioral study of the post-axotomy functionality, (4) the sample population is well contained and experiment conditions are easily reproducible since the trap for surgery and the environment for recovery are on the same chip, and finally (5) the design of the chip is simple enough to be adapted to other organisms or many other kinds of experiments, including: ablation, irradiation, stimulation or simply observation, widening the possibilities of high-throughput biological investigations.
Supplementary Material
Supplementary Figure 1 Schematic of the femtosecond laser nanoaxotomy setup.
Supplementary Figure 2 Nanoaxotomy on-a-chip.
Supplementary Figure 3 Locations of nanoaxotomy in PLM process.
Supplementary Table 1 Sequence of operations.
Supplementary Table 2 Summary of nanoaxotomy experiments performed at different locations.
Supplementary Methods
Supplementary Video 1 Immobilization of a worm in the microfluidic trap.
Supplementary Video 2 Laser nanoaxotomy of a worm on a chip.
Supplementary Video 3 Behavioral assay of a worm following nanoaxotomy on the branches of all of its PLM and ALM processes.
Acknowledgments
The authors would like to acknowledge Prof. Cornelia Bargmann and Prof. Miriam Goodman for their valuable discussions. This work was supported by grants NS058646 and NS060129 from the National Institute of Health and by a grant BES-0548673 from the National Science Foundation.
Footnotes
AUTHOR CONTRIBUTIONS
N.C and A.B. designed the device; T.C. fabricated the microfluidic chip; S.X.G. and N.D. performed two-photon microscopy; F.B., M.H., and A.B. analyzed data, S.X.G., F.B., and A.B. designed experiments, S.X.G. and F.B. performed experiments, F.B. and A.B. wrote the paper.
Note: Supplementary information is available on the Nature Methods website.
References
- 1.Yanik MF, Cinar H, Cinar HN, Chisholm AD, Jin Y, Ben-Yakar A. Nature. 2004;432:882. doi: 10.1038/432822a. [DOI] [PubMed] [Google Scholar]
- 2.Bourgeois F, Ben-Yakar A. Opt Exp. 2007;15(14):8521–8531. doi: 10.1364/oe.15.008521. [DOI] [PubMed] [Google Scholar]
- 3.Gray JM, Karow DS, Lu H, Chang AJ, Chang JS, Ellis RE, Marietta MA, Bargmann CI. Nature. 2005;430:317–322. doi: 10.1038/nature02714. [DOI] [PubMed] [Google Scholar]
- 4.Chronis N, Zimmer M, Bargmann CI. Nat Meth. 2007;4(9):729–731. doi: 10.1038/nmeth1075. [DOI] [PubMed] [Google Scholar]
- 5.Chalasani SH, Chronis N, Tsunozaki M, Gray JM, Ramot D, Goodman MB, Bargmann CI. Nature. 2007;450:63–70. doi: 10.1038/nature06292. [DOI] [PubMed] [Google Scholar]
- 6.Hulme SE, Shevkoplyas SS, Apfeld J, Fontana W, Whitesides GM. Lab Chip. 2007;7(11):1515–1523. doi: 10.1039/b707861g. [DOI] [PubMed] [Google Scholar]
- 7.Kima N, Dempsey CM, Zoval JV, Sze JY, Madoub MJ. Sens Actuat B. 2007;122:511–518. [Google Scholar]
- 8.Lange D, Storment CW, Conley CA, Kovacs GTA. Sens Actuat B. 2005;107:904–914. [Google Scholar]
- 9.Qin J, Wheeler AR. Lab Chip. 2007;7(2):186–192. doi: 10.1039/b613414a. [DOI] [PubMed] [Google Scholar]
- 10.Park SJ, Goodman MB, Pruitt BL. PNAS. 2007;104(44):17376–17381. doi: 10.1073/pnas.0702138104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rohde CB, Zeng F, Gonzalez-Rubio R, Angel M, Yanik MF. PNAS. 2007;104(35):13891–13895. doi: 10.1073/pnas.0706513104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Unger MA, Chou HP, Thorsen T, Scherer A, Quake SR. Science. 2000;288:113–116. doi: 10.1126/science.288.5463.113. [DOI] [PubMed] [Google Scholar]
- 13.Kandel ER, Schwartz JH, Jessell TM. Principles of neural science. 4. McGraw-Hill Medical; New York, NY, USA: 2000. [Google Scholar]
- 14.Chalfie M, Sulston JE, White JG, Southgate E, Thomson JN, Brenner S. J Neurosci. 1985;5:956–964. doi: 10.1523/JNEUROSCI.05-04-00956.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu Z, Ghosh-Roy A, Yanik MF, Zhang JZ, Jin Y, Chisholm AD. PNAS. 2007;104:15132. doi: 10.1073/pnas.0707001104. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure 1 Schematic of the femtosecond laser nanoaxotomy setup.
Supplementary Figure 2 Nanoaxotomy on-a-chip.
Supplementary Figure 3 Locations of nanoaxotomy in PLM process.
Supplementary Table 1 Sequence of operations.
Supplementary Table 2 Summary of nanoaxotomy experiments performed at different locations.
Supplementary Methods
Supplementary Video 1 Immobilization of a worm in the microfluidic trap.
Supplementary Video 2 Laser nanoaxotomy of a worm on a chip.
Supplementary Video 3 Behavioral assay of a worm following nanoaxotomy on the branches of all of its PLM and ALM processes.