

Abstract
Heat shock protein 90 (Hsp90) is a molecular chaperone whose association is required for the stability and function of multiple mutated, chimeric and over‐expressed signaling proteins that promote the growth and/or survival of cancer cells. Hsp90 client proteins include mutated p53, Bcr‐Abl, Raf‐1, Akt, HER2/Neu (ErbB2) and HIF‐1α. Hsp90 inhibitors, by interacting specifically with a single molecular target, cause the destabilization and eventual degradation of Hsp90 client proteins, and the first‐in‐class Hsp90 inhibitor, 17‐allylamino‐17 demethoxygeldanamycin (17AAG), is currently in phase II clinical trials. A fraction of Hsp90 has been identified at the cell surface and its presence has recently been shown to correlate with melanoma progression. Inhibition of cell‐surface Hsp90 with antibodies or cell‐impermeable Hsp90 inhibitors blocks cell motility and invasion in vitro and cancer metastasis in vivo. Thus, cell‐surface Hsp90 may play a unique role in tumor metastasis, distinct from but perhaps overlapping with its intracellular function. In addition, because cell‐surface Hsp90 may be the point of contact between some viruses and host cells, this pool of the chaperone may play a distinct role in initiation of infectious disease. (Cancer Sci 2007; 98: 1536–1539)
- Abbreviations: 17AAG
17‐allylamino‐17 demethoxygeldanamycin
- 17DMAG
17‐dimethylaminoethylamino‐17‐demethoxygeldanamycin
- APC
antigen‐presenting cells
- DEN
Dengue
- DMAG‐N‐oxide
17‐(2‐Dimethyloxidoaminoethyl) amino‐17‐demethoxygeldanamycin
- FAK
focal adhesion kinase
- GA
geldanamycin
- Hsp90
heat shock protein 90
- LPS
lipopolysaccharide
- MHC
major histocompatibility complex
- MMP
matrix metalloproteases
- NF‐κB
nuclear factor‐κB
- NK
natural killer
- TLR4
Toll‐like receptor 4.
Hsp90 as a target for cancer therapy
In the post‐genomic era, identification of novel molecular targets for cancer therapeutics offers the promise of great specificity coupled with reduced systemic toxicity, but simultaneously faces the potential peril of being unable to deal successfully with diseases that are frequently caused by multiple genetic abnormalities (e.g. cancer). Indeed, a pessimistic viewpoint might hold that the more finely tuned our molecular therapies become, the less they will be able to confront effectively the inherent genetic instability of cancer cells. Partly because of their genetic plasticity, cancer cells are very efficient at adapting to a noxious environment, whether that is hypoxia, deprivation of certain hormones or growth factors, or exposure to chemotherapy or irradiation. Thus, hormone‐dependent tumor cells eventually become hormone‐independent, either by receptor mutation or by other means. Likewise, tumor cells exposed to hypoxia induce a multifaceted transcriptional response geared to survive such an environment. Similarly, cancer cells exposed to levels of chemotherapy that are initially fatal eventually activate multiple and overlapping signaling pathways to protect themselves from further harm.
One approach to this dilemma might be to target the very machinery that allows cancer cells to adapt so successfully to environmental stress. Cells respond to environmental stress by increasing synthesis of several molecular chaperones (also known as Hsp, because they were first observed in cells exposed to elevated temperature). These housekeeping proteins, as their name implies, assist general protein folding and prevent non‐functional side reactions such as the non‐specific aggregation of misfolded or unfolded proteins.
Hsp90 is one of the most abundant molecular chaperones whose association is important for maintaining the stability and function of numerous client proteins.( 1 ) In many cases, Hsp90 client proteins are mutated or activated in cancer cells. For example, Hsp90 had been found in complex with the oncogenic tyrosine kinase v‐Src, the mutated oncogene Bcr/Abl, and the serine/threonine kinase Raf‐1.( 2 ) Hsp90 is constitutively expressed at 2–10‐fold higher levels in tumor cells, comprising 1–2% of cell protein, compared with their normal counterparts( 3 ) suggesting that it could be crucially important for the growth and/or survival of tumor cells. Indeed, Hsp90 over‐expression in breast cancer cells has been correlated with acquired resistance to some forms of chemotherapy.( 4 ) Furthermore, Kamal et al. reported that Hsp90 proteins are activated in tumor cells and facilitate malignant progression.( 5 ) In short, accumulating evidence suggests that the chaperone activity of Hsp90 may be crucial for signaling proteins that contribute to cancer progression.
In 1994, Whitesell et al. reported that a class of antibiotics known as benzoquinone ansamycins, and GA in particular, bound specifically to Hsp90, inhibited the association of the chaperone with v‐Src protein, and led to the eventual destabilization of the protein.( 6 ) Although GA itself proved to be too hepatotoxic for clinical use, a better tolerated derivative (17AAG) that also binds Hsp90 has shown promising anti‐tumor activity and predicted biological activity in preclinical models, and is now in multiple phase II clinical trials.( 7 ) Further, additional structurally unique Hsp90 inhibitors are undergoing preclinical and clinical evaluation.
Extracellular Hsp90
While intracellular Hsp90 has thus become a very exciting anticancer molecular target, a relatively small number of studies have suggested that a pool of the chaperone is located on the cell surface, facing the extracellular space, and that this fraction of Hsp90 may have unique and unexpected properties. As shown in Table 1, Hsp90 has been identified at the surface of a number of cell types. As long ago as 1986, Ullrich et al. identified a cell surface tumor‐specific transplantation antigen that they identified as Hsp90.( 8 ) Immunization of mice with this antigen significantly inhibited subsequent tumor growth, suggesting that extracellular Hsp90 must contribute to the host anti‐tumor immune response. Recently, Hsp90 expression was reported to be dramatically up‐regulated in malignant melanoma cells when compared to benign melanocytic lesions, and Hsp90 was expressed on the surface of seven of eight melanoma metastases.( 9 ) Cancer‐specific expression of cell‐surface Hsp90 has also been observed in small‐cell lung cancer cells, and the protein was found to be associated with MHC class I.( 3 ) Lymphoma cells have also been reported to express Hsp90 at their cell surface, with the expression level increasing through several stages of early and late apoptotic death in conjunction with immune response activation.( 10 ) Finally, the proteasome inhibitor Bortezomib induces cell‐surface expression of Hsp90 in dying melanoma cells, with concomitant stimulation of dendritic cell activity and enhancement of anti‐tumor immunity.( 11 ) Thus, accumulating evidence indicates that extracellular Hsp90 might be a useful tumor antigen for eliciting a host immune response. The reader is referred to a recent review summarizing this strategy.( 12 )
Table 1.
Possible roles of extracellular heat shock protein (Hsp)90
| Cell type | Function | Reference |
|---|---|---|
| Lung cancer, melanoma | Immune response | ( 3 ) |
| Hepatoma | ||
| Fibrosarcoma | Immune response | ( 8 ) |
| Lymphoma | Immune response | ( 10 ) |
| Macrophage, dendritic cell | Immune response | ( 19 ) |
| Monocyte | Immune response | ( 20 ) |
| Melanoma | Metastasis | ( 9 ) |
| Fibrosarcoma | Invasion | ( 13 ) |
| Neuronal cell | Migration | ( 14 ) |
| Melanoma | Invasion/migration | ( 15 ) |
| Dermal fibroblast | Migration | ( 16 ) |
Cell‐surface Hsp90 and cancer metastasis
In 2004, Eustace et al. identified cell‐surface Hsp90 to be crucial for the invasiveness of HT‐1080 fibrosarcoma cells in vitro. ( 13 ) In the Eustace et al. study, the investigators demonstrated that the pharmacologic Hsp90 inhibitor GA, covalently affixed to cell‐impermeable beads, was able to significantly impair cell invasion. Coincident with its inhibitory effect on cell invasiveness, cell‐impermeable GA also antagonized the maturation, via proteolytic self‐processing, of MMP2, a cell‐surface enzyme whose activity has been previously demonstrated to be essential for cell invasion. Furthermore, these investigators demonstrated that Hsp90 could be found in association with MMP2 in the culture medium bathing the HT‐1080 cells. It is intriguing to speculate that association with Hsp90 on the cell surface is necessary for the self‐proteolysis of MMP2.
In the same year (2004), Sidera et al. reported that cell‐surface Hsp90 is crucial for cell motility in neuronal cells.( 14 ) Neutralizing antibodies against cell surface Hsp90 inhibited Schwann cell migration. In fact, Hsp90 antibodies inhibited actin reorganization, which is known to be an important component of cell migration. These investigators showed that a neutralizing Hsp90 antibody inhibited not only neuronal cell migration but also melanoma migration.( 15 ) Furthermore, treatment of mice with Hsp90 antibody inhibited melanoma metastasis in vivo.
Very recently, Li et al. reported that extracellular Hsp90 regulates cell motility in human dermal fibroblast cells.( 16 ) They demonstrated that treatment of neutralizing antibodies against cell‐surface Hsp90 inhibited cell motility, and that treatment of recombinant Hsp90 protein stimulated cell motility in vitro. Furthermore, treatment of mice with cream that contained recombinant Hsp90 protein enhanced wound healing in vivo. Interestingly, hypoxia stimulated secretion of Hsp90 through induction of hypoxia inducible factor (HIF)‐1α expression.
These findings have been confirmed using a cell‐impermeable polar derivative of 17DMAG, a small molecule Hsp90 inhibitor structurally related to GA and currently in clinical trial (S. Tsutsumi, unpublished data, 2007). In analyzing a number of GA‐derived Hsp90 inhibitors for their membrane permeability and affinity for Hsp90, the authors identified DMAG‐N‐oxide as a potent cell‐impermeable Hsp90 inhibitor. Although it lacks the well‐recognized growth inhibitory capability of cell‐permeable Hsp90 inhibitors, it displays anti‐invasive activity in vitro and anti‐metastatic activity in vivo. DMAG‐N‐oxide inhibited cell invasion in bladder cancer, prostate cancer and melanoma models. DMAG‐N‐oxide inhibited serum‐dependent cell migration and actin re‐organization. Interestingly, DMAG‐N‐oxide inhibited both matrix‐dependent integrin–Src interaction and FAK–Src interaction, suggesting that cell‐surface Hsp90 might regulate focal adhesion formation through modulation of integrin signaling. In an experimental murine melanoma model, in vivo administration of cell‐impermeable Hsp90 inhibitors reduced the frequency of metastasis and significantly improved survival.
Extracellular Hsp90 interacting proteins
As summarized in Fig. 1, although more than 100 intracellular Hsp90 client proteins have been identified, much less is known about extracellular Hsp90 client proteins. As mentioned above, extracellular Hsp90 is reported to bind to MMP‐2 and its association is necessary to generate the mature enzyme.( 13 ) Very recently, cell‐surface Hsp90 was reported to interact with the extracellular domain of ErbB2,( 17 ) a protein whose intracellular kinase domain is already known to interact with Hsp90.( 18 ) Additional cell‐surface Hsp90‐binding proteins have been identified. Extracellular Hsp90 has been shown to be taken up by APC and re‐presented by MHC class I molecules. When added to intact cells, Hsp90 interacts with CD91, suggesting that CD91 acts as a Hsp90 receptor.( 19 ) TLR4 is another Hsp90 receptor. Cell surface Hsp90 clusters with TLR4 within lipid rafts following LPS stimulation.( 20 ) These Hsp90 receptors appear to be crucial for the activity of NK cells and APC such as a macrophages and dendritic cells.( 12 ) Annexin II is another extracellular Hsp90 binding protein.( 21 ) High glucose increases the expression of cell‐surface Hsp90 in aortic endothelial cells, and promotes the association of Hsp90 with annexin II, which enhances the generation of plasmin. Furthermore, cell‐surface Hsp90 is elevated in the aortas of diabetic rats, suggesting that cell‐surface Hsp90 may contribute to clotting abnormalities associated with the diabetic state.
Figure 1.
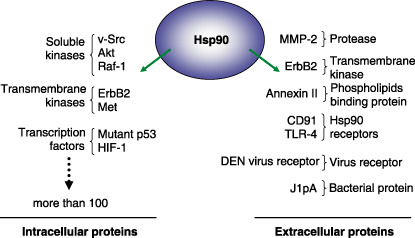
A partial list of intracellular and extracellular heat shock protein (Hsp)90 client proteins.
By interacting with various viral and bacterial proteins, extracellular Hsp90 may also play an important role in infectious disease. Campylobacter jejuni is a leading cause of acute bacterial gastroenteritis in humans. C. jejuni surface lipoprotein, JlpA, is important for adherence of the bacterium to epithelial cells. JlpA interacts with cell‐surface Hsp90 and regulates the activity of NF‐κB and p38 MAP kinase in host cells. GA and Hsp90 antibody inhibit the interaction of JlpA with Hsp90, suggesting that cell‐surface Hsp90 may be capable of mediating bacterial infection.( 22 )
DEN virus is the most important arthropod‐borne human pathogen and it is transmitted to humans by the bite of the mosquito. The first step in DEN virus infection requires interaction between the DEN E protein (envelope glycoprotein) and a receptor present on the surface of the host cell. Cell‐surface Hsp90 has been identified as a component of the DEN virus receptor complex, and treatment of host cells with Hsp90 antibody has been shown to inhibit DEN virus infection. Taken together, these data suggest that cell‐surface Hsp90 is a potentially important target of novel drugs to combat viral infection.( 23 )
Other cell‐surface Hsp and cancer progression
As summarized in Table 2, it has been shown that not only Hsp90 but also other Hsp are expressed on the cell surface and that these proteins are crucial for cancer progression. Hsp25, a murine homolog of human Hsp27, has been shown to regulate metastasis.( 24 ) Surface expression of Hsp25 correlates with enhanced breast cancer growth in vivo, and cells expressing higher levels of Hsp25 on the cell surface display enhanced metastatic potential in vitro and enhanced metastatic activity in vivo. Hsp70, a major stress‐inducible chaperone, is expressed at the cell surface of melanoma metastases but not in normal skin fibroblasts.( 25 ) Cell‐surface expression of Hsp70 is observed to be significantly higher in clinically advanced oral tumors,( 26 ) suggesting that cell‐surface Hsp70 might be crucial for tumor progression. In contrast, cell‐surface Hsp70 seems to inhibit metastasis in breast cancer cells.( 24 ) This discrepancy may be due to the difference in cell type. Gp96, the Hsp90 homolog in endoplasmic reticulum, has been shown to be expressed at the cell surface only in tumor cells,( 27 ) and it was the first chaperone described to play a role in the induction of anti‐tumor immune responses. Several phase I and phase II clinical trials are currently ongoing using a Gp96‐peptide complex as a cancer vaccine. The role of Gp96 in mediating immune responses was well summarized in a recent review.( 28 )
Table 2.
Possible roles of extracellular heat shock proteins in cancer metastasis
Calreticulin, an endoplasmic reticulum chaperone, is also expressed at the cell surface. Calreticulin has been identified as the cell surface lectin responsible for triggering cell spreading of melanoma cells.( 29 ) Treatment of cells with calreticulin antibody inhibited laminin‐dependent cell spreading, suggesting that calreticulin is crucial for cell motility and that it might participate in integrin signaling. Indeed, it is reported that calreticulin is associated with integrin α‐2/β‐1 on the platelet surface, and that it modulates ligand interaction with integrins.( 30 )
Interestingly, Hsp47, which is known as a rheumatoid arthritis‐related antigen, has been shown to be expressed at the cell surface, and oral cancer cells expressing high levels of surface Hsp47 display low invasive activity, suggesting that Hsp47 has an inhibitory effect on cell migration/invasion.( 31 ) Thus, many distinct chaperone proteins might be expressed at the cell surface, and have diverse effects on cell motility, invasion, and cancer progression.
Conclusion
Accumulating evidence supports the continued development of Hsp90 inhibitors that specifically target the cell surface pool of this chaperone. Such agents may have benefit not only in the cancer metastasis paradigm but also as a possible strategy to prevent viral infection. It may also be useful to inhibit (or stimulate) other Hsp as a novel therapeutic strategy specifically targeting cancer metastasis. However, questions remain concerning the mechanism of action of these chaperones in general and Hsp90 in particular: (i) how are Hsp90 and the other chaperones secreted?; and (ii) does extracellular Hsp90 depend on its ATPase activity or association with other co‐chaperones for its function? Because Hsp90 does not have a traditional secretory sequence, it is likely secreted by non‐traditional mechanisms. Hsp70, which also lacks a secretory sequence, is known to be secreted by a non‐traditional pathway that depends on exosomes.( 32 ) Indeed, Hsp90 is known to be present in exosomes,( 33 ) suggesting that Hsp90 might also be secreted by an exosome pathway. Previous reports have identified the Hsp90 co‐chaperones, p48Hip and p60Hop, to be secreted by fibrosarcoma cells,( 34 ) and extracellular Hsp90 has been reported to form complexes with other chaperone proteins at cell‐surface receptors,( 20 , 23 ) although it remains possible that Hsp90 may not require additional proteins to subserve its cell surface function.
Other questions also await further experimentation. For example, what effect (if any) does surface Hsp90 inhibition have on the generation of tumor‐specific immunity by the host? Is the tumor‐specific immunity that is mediated by other surface‐expressed chaperone proteins affected? Is Hsp90 inhibition with antibody equivalent to its inhibition by a small molecule? While Hsp90 and other molecular chaperones are well‐recognized to play important roles in regulating intracellular homeostasis, distinct extracellular pools of these proteins appear to have unique, previously unrecognized but equally important activities. Further investigation is needed to provide answers to the difficult questions raised by these preliminary observations.
References
- 1. Neckers L, Neckers K. Heat‐shock protein 90 inhibitors as novel cancer chemotherapeutics – an update. Expert Opin Emerg Drugs 2005; 10: 137–49. [DOI] [PubMed] [Google Scholar]
- 2. Neckers L. Hsp90 inhibitors as novel cancer chemotherapeutic agents. Trends Mol Med 2002; 8: S55–61. [DOI] [PubMed] [Google Scholar]
- 3. Ferrarini M, Heltai S, Zocchi MR, Rugarli C. Unusual expression and localization of heat‐shock proteins in human tumor cells. Int J Cancer 1992; 51: 613–19. [DOI] [PubMed] [Google Scholar]
- 4. Beliakoff J, Whitesell L. Hsp90: an emerging target for breast cancer therapy. Anticancer Drugs 2004; 15: 651–62. [DOI] [PubMed] [Google Scholar]
- 5. Kamal A, Thao L, Sensintaffar J et al . A high‐affinity conformation of Hsp90 confers tumour selectivity on Hsp90 inhibitors. Nature 2003; 425: 407–10. [DOI] [PubMed] [Google Scholar]
- 6. Whitesell L, Mimnaugh EG, De Costa B, Myers CE, Neckers LM. Inhibition of heat shock protein HSP90‐pp60v‐src heteroprotein complex formation by benzoquinone ansamycins: essential role for stress proteins in oncogenic transformation. Proc Natl Acad Sci USA 1994; 91: 8324–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Sharp S, Workman P. Inhibitors of the HSP90 molecular chaperone: current status. Adv Cancer Res 2006; 95: 323–48. [DOI] [PubMed] [Google Scholar]
- 8. Ullrich SJ, Robinson EA, Law LW, Willingham M, Appella E. A mouse tumor‐specific transplantation antigen is a heat shock‐related protein. Proc Natl Acad Sci USA 1986; 83: 3121–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Becker B, Multhoff G, Farkas B et al . Induction of Hsp90 protein expression in malignant melanomas and melanoma metastases. Exp Dermatol 2004; 13: 27–32. [DOI] [PubMed] [Google Scholar]
- 10. Sapozhnikov AM, Ponomarev ED, Tarasenko TN, Telford WG. Spontaneous apoptosis and expression of cell surface heat‐shock proteins in cultured EL‐4 lymphoma cells. Cell Prolif 1999; 32: 363–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Spisek R, Charalambous A, Mazumder A, Vesole DH, Jagannath S, Dhodapkar MV. Bortezomib enhances dendritic cell (DC) mediated induction of immunity to human myeloma via exposure of cell surface heat shock protein 90 on dying tumor cells: therapeutic implications. Blood 2007; 109: 4839–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Binder RJ, Vatner R, Srivastava P. The heat‐shock protein receptors: some answers and more questions. Tissue Antigens 2004; 64: 442–51. [DOI] [PubMed] [Google Scholar]
- 13. Eustace BK, Sakurai T, Stewart JK et al . Functional proteomic screens reveal an essential extracellular role for hsp90 alpha in cancer cell invasiveness. Nat Cell Biol 2004; 6: 507–14. [DOI] [PubMed] [Google Scholar]
- 14. Sidera K, Samiotaki M, Yfanti E, Panayotou G, Patsavoudi E. Involvement of cell surface HSP90 in cell migration reveals a novel role in the developing nervous system. J Biol Chem 2004; 279: 45 379–88. [DOI] [PubMed] [Google Scholar]
- 15. Stellas D, Karameris A, Patsavoudi E. Monoclonal antibody 4C5 immunostains human melanomas and inhibits melanoma cell invasion and metastasis. Clin Cancer Res 2007; 13: 1831–8. [DOI] [PubMed] [Google Scholar]
- 16. Li W, Li Y, Guan S et al . Extracellular heat shock protein‐90alpha: linking hypoxia to skin cell motility and wound healing. Embo J 2007; 26: 1221–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Sidera K, Gaitanou M, Matsas R, Patsavoudi E. Cell surface hsp90 interacts with the extracellular domain of her2 and contributes to receptor activation. FEBS J 2006; 273: 95. [Google Scholar]
- 18. Xu W, Yuan X, Xiang Z, Mimnaugh E, Marcu M, Neckers L. Surface charge and hydrophobicity determine ErbB2 binding to the Hsp90 chaperone complex. Nat Struct Mol Biol 2005; 12: 120–6. [DOI] [PubMed] [Google Scholar]
- 19. Basu S, Binder RJ, Ramalingam T, Srivastava PK. CD91 is a common receptor for heat shock proteins gp96, hsp90, hsp70, and calreticulin. Immunity 2001; 14: 303–13. [DOI] [PubMed] [Google Scholar]
- 20. Triantafilou M, Triantafilou K. Heat‐shock protein 70 and heat‐shock protein 90 associate with toll‐like receptor 4 in response to bacterial lipopolysaccharide. Biochem Soc Trans 2004; 32: 636–9. [DOI] [PubMed] [Google Scholar]
- 21. Lei H, Romeo G, Kazlauskas A. Heat shock protein 90alpha‐dependent translocation of annexin II to the surface of endothelial cells modulates plasmin activity in the diabetic rat aorta. Circ Res 2004; 94: 902–9. [DOI] [PubMed] [Google Scholar]
- 22. Jin S, Song YC, Emili A, Sherman PM, Chan VL. JlpA of Campylobacter jejuni interacts with surface‐exposed heat shock protein 90alpha and triggers signalling pathways leading to the activation of NF‐kappaB and p38 MAP kinase in epithelial cells. Cell Microbiol 2003; 5: 165–74. [DOI] [PubMed] [Google Scholar]
- 23. Reyes‐Del Valle J, Chavez‐Salinas S, Medina F, Del Angel RM. Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol 2005; 79: 4557–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Bausero MA, Page DT, Osinaga E, Asea A. Surface expression of Hsp25 and Hsp72 differentially regulates tumor growth and metastasis. Tumour Biol 2004; 25: 243–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Farkas B, Hantschel M, Magyarlaki M et al . Heat shock protein 70 membrane expression and melanoma‐associated marker phenotype in primary and metastatic melanoma. Melanoma Res 2003; 13: 147–52. [DOI] [PubMed] [Google Scholar]
- 26. Kaur J, Das SN, Srivastava A, Ralhan R. Cell surface expression of 70 kDa heat shock protein in human oral dysplasia and squamous cell carcinoma: correlation with clinicopathological features. Oral Oncol 1998; 34: 93–8. [DOI] [PubMed] [Google Scholar]
- 27. Altmeyer A, Maki RG, Feldweg AM et al . Tumor‐specific cell surface expression of the‐KDEL containing, endoplasmic reticular heat shock protein gp96. Int J Cancer 1996; 69: 340–9. [DOI] [PubMed] [Google Scholar]
- 28. Li Z, Dai J, Zheng H, Liu B, Caudill M. An integrated view of the roles and mechanisms of heat shock protein gp96‐peptide complex in eliciting immune response. Front Biosci 2002; 7: d731–51. [DOI] [PubMed] [Google Scholar]
- 29. White TK, Zhu Q, Tanzer ML. Cell surface calreticulin is a putative mannoside lectin which triggers mouse melanoma cell spreading. J Biol Chem 1995; 270: 15 926–9. [DOI] [PubMed] [Google Scholar]
- 30. Elton CM, Smethurst PA, Eggleton P, Farndale RW. Physical and functional interaction between cell‐surface calreticulin and the collagen receptors integrin alpha2beta1 and glycoprotein VI in human platelets. Thromb Haemost 2002; 88: 648–54. [PubMed] [Google Scholar]
- 31. Hebert C, Norris K, Della Coletta R, Reynolds M, Ordonez J, Sauk JJ. Cell surface colligin/Hsp47 associates with tetraspanin protein CD9 in epidermoid carcinoma cell lines. J Cell Biochem 1999; 73: 248–58. [DOI] [PubMed] [Google Scholar]
- 32. Lancaster GI, Febbraio MA. Exosome‐dependent trafficking of HSP70: a novel secretory pathway for cellular stress proteins. J Biol Chem 2005; 280: 23 349–55. [DOI] [PubMed] [Google Scholar]
- 33. Clayton A, Turkes A, Navabi H, Mason MD, Tabi Z. Induction of heat shock proteins in B‐cell exosomes. J Cell Sci 2005; 118: 3631–8. [DOI] [PubMed] [Google Scholar]
- 34. Eustace BK, Jay DG. Extracellular roles for the molecular chaperone, hsp90. Cell Cycle 2004; 3: 1098–100. [PubMed] [Google Scholar]